- Osteichthyes
-
 Peces óseos
Peces óseosRango temporal: Devónico - Presente 
Epinephelus lanceolatusClasificación científica Reino: Animalia Filo: Chordata Subfilo: Vertebrata Superclase: Gnathostomata Clases Los osteictios (Osteichthyes, del griego ὀστεον /ósteon/, "hueso", e ἰχθύς /ichtus/ , "pez") o peces óseos son vertebrados gnatóstomos que incluye a todos los peces dotados de esqueleto interno óseo, es decir, hecho principalmente de piezas calcificadas y muy pocas de cartílago. Junto con los condrictios (peces cartilaginosos), forman los dos grandes grupos que comprenden los animales llamados vulgarmente «peces»; aún existe un tercer grupo, el de los peces sin mandíbulas (lampreas y mixines), con muy pocos representantes actuales. El término Osteichthyes hace referencia a un grupo parafilético por lo que no es una clasificación válida. Hoy en día se sabe que este grupo está formado por las clases Actinopterygii y Sarcopterygii.
Contenido
Clasificación
Los peces óseos comprenden dos clases:[1] [2]
- Clase Actinopterygii
- Subclase Chondrostei
- Subclase Neopterygii
- Clase Sarcopterygii
- Subclase Coelacanthimorpha
- Subclase Dipnoi
- Subclase Tetrapodomorpha (tetrápodos y sus ancestros pisciformes directos)
Los actinopterigios comprenden la inmensa mayoría de los peces, caracterizados por una estructura ósea radial en las aletas. Los sarcopterigios comprenden los peces con aletas pares lobuladas, es decir, los celacantos y peces pulmonados, que están directamente emparentados con los vertebrados terrestres (Tetrapoda). Por ello, y según la sistemática cladística, los tetrápodos se incluyen en esta clase.
Filogenia
En términos cladísticos, los osteictios son un grupo parafilético, es decir, las dos subclases que incluye, actinopterigios y sarcopterigios no muestran ningún carácter derivado en común; de hecho, los sarcopterigios son más cercanos a los tetrápodos (anfibios y demás vertebrados con cuatro patas) que a los actinopterigios, como se puede comprobar en el siguiente cladograma:[3]
Gnathostomata Chondrichthyes (peces cartilaginosos)
Teleostomi Actinopterygii (peces óseos con aletas con radios)
Sarcopterygii (peces óseos con aletas lobuladas)
Tetrapoda (vertebrados con cuatro patas)
Características
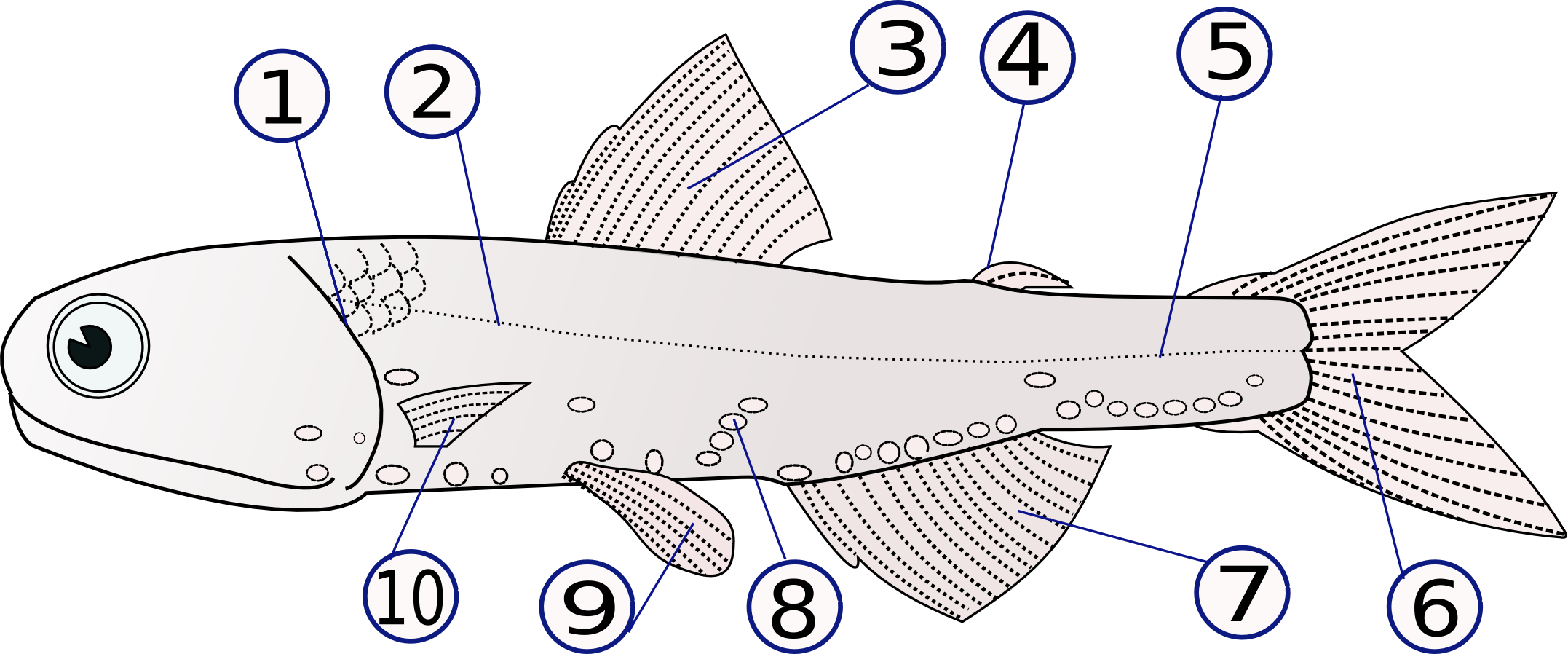
 Lampanyctodes hectoris
Lampanyctodes hectoris
(1) Opérculo, (2) Línea lateral, (3) Aleta dorsal, (4) Aleta adiposa, (5) Pedúnculo caudal, (6) Aleta caudal, (7) Aleta anal, (8) Fotóforos, (9) Aletas pelvianas (par), (10) Aletas torácicas (par)Internamente se distinguen algunas diferencias con los peces cartilaginosos: mientras que éstos poseen una válvula espiral en el intestino, los osteictios presentan ciegos pilóricos y carecen de glándula rectal. En el Sistema respiratorio aparecen branquias dentro de una cámara branquial, y recubiertas por un opérculo que muestra al exterior una sola abertura branquial a cada lado. Ocasionalmente puede marcarse un preopérculo. Además, las branquias no están separadas por septos. En algunos grupos, la vejiga natatoria se ha transformado en un pulmón original, que le sirve además para flotar a determinado nivel, o desplazarse verticalmente. Ambas estructuras, pulmón y vejiga natatoria, son mutuamente excluyentes, es decir, cuando existe uno no existe el otro, y viceversa.
Presentan mayoritariamente boca terminal, capaz de realizar movimientos muy precisos gracias a que poseen huesos dérmicos articulados. Los dientes salen de algunos de estos huesos dérmicos y carecen de reemplazo cuando se caen o rompen. Además, su aleta caudal es homocerca. Aparte de su esqueleto interno calcificado u osificado (la excepción es el esturión, que lo tiene cartilaginoso como los condrictios), también poseen huesos en la dermis de su tegumento en forma de escamas, lo que se conoce como dermatoesqueleto. Estas escamas tienen un importante valor taxonómico, ya que el tipo y número (especialmente en la línea lateral y transversalmente) son usados como rasgos identificativos de los distintos grupos.
En relación a las aletas de los osteíctios, encontramos mayoritariamente un par de aletas pelvianas y un par de aletas torácicas o pectorales (ambas aletas son pares y simétricas en el cuerpo) y una o varias dorsales o una o varias anales en el plano sagital, pero no de forma simétrica. También presentan aleta caudal.
Dependiendo de la posición de las aletas pelvianas y torácicas, podemos distinguir cuatro tipos de osteíctios: abdominales (si las aletas pelvianas se encuentran situadas por detrás de las torácicas), torácicos (si están a la misma altura o ligeramente retrasadas), yugulares (si están por delante) o ápodos (carecen de aletas pelvianas).
-

Abdominal. Lucio europeo (Esox lucius)
-

Torácico. (Plectorhinchus vittatus)
-

Yugular. Pez araña (Trachinus draco)
-

Ápodo. Anguila europea (Anguilla anguilla)
Algunas especies son gregarias y se reúnen en bancos por medio de señales acústicas. Son, en general, animales carnívoros, aunque también existen especies planctónicas, e incluso herbívoros. El nivel de adaptación al medio hace de los peces óseos los más numerosos de todos los grupos.
Locomoción
La mayoría de los osteíctios poseen vejiga natatoria que les permite ser ingrávidos en el agua, ya que la llenan o vacían de gas para controlar la profundidad en la que se encuentran. Aprovechan mejor el movimiento de ascenso o descenso pegando las aletas al cuerpo y evitando el rozamiento. Las aletas se conservan, no obstante, porque resultan útiles para movimientos más precisos. Además, las aletas han evolucionado originando distintas funciones, como servir para caminar por el fondo (Periophthalmus koelreuteri), para palpar (Tríglidos, Trigla), como ventosas (Lepadogaster), o como alas para saltar durante un tiempo por encima de la superficie (Cheilopogon) evitando a los depredadores acuáticos en situaciones de estrés.
La morfología típica de los peces es la que menos resistencia hidrodinámica ofrece para la natación: el extremo anterior aguzado y una altura igual a un cuarto de la longitud del cuerpo, estando la altura máxima en el primer tercio del cuerpo contando desde la cabeza. Esta forma permite minimizar las turbulencias facilitando el avance.
De todos modos, no todos los peces presentan una morfología similar a la descrita. La longitud del animal varía de unos grupos a otros, así como la parte propulsora del movimiento. Los anguiliformes tienen una parte propulsora muy alargada; los carangiformes son intermedios y los más típicos; y los ostraciformes tienen una parte propulsora muy pequeña. Estas diferentes formas responden a diferentes y variadas formas de vida de los peces.
Dentro de los carangiformes destaca un grupo especial, los escombriformes, que es el tipo de movimiento mejor adaptado a una natación continua. Lo presentan atunes, bonitos, caballas. Tienen un cuerpo muy musculoso con muy baja resistencia al agua: la cola es lo más delgada posible, semilunar y rígida, y tienen pínnulas que sirven para reducir las turbulencias. Son por tanto muy hidrodinámicos, perfeccionando este rasgo con un rostro en punta y con unos surcos que permiten a las aletas pares e impares retraerse y no ofrecer resistencia al movimiento. Para poder tener una elevada actividad muscular, mantienen la temperatura corporal por encima de la del medio, por lo que se dice que son pseudohomeotermos. Los individuos grandes mantienen velocidades de 60 a 70 km/h, y superan en velocidad punta los 100 km/h.
Anatomía
Tegumento
El tegumento, o piel, está formado por la dermis y epidermis.
Epidermis y dermis
La dermis de los osteíctios presenta escamas imbricadas (como las tejas de una casa), y sobre las escamas se dispone la epidermis. Ésta posee unas diez capas de células no queratinizadas y vivas (a diferencia del resto de los tetrápodos) y una capa basal, que es la que se va dividiendo y originando nuevas células epidérmicas.
La epidermis presenta muchas células mucosas (glándulas unicelulares que secretan mucus). Este mucus regula la permeabilidad de la piel (y por tanto la excreción) y reduce la fricción entre las escamas, permitiendo que éstas se articulen correctamente. El mucus también sirve para lubricar el cuerpo y para defenderlo de patógenos.
La dermis, más gruesa, contiene escamas, cromatóforos (células responsables de la coloración de los peces), tejido conjuntivo, vasos sanguíneos y terminaciones nerviosas.
Escamas dérmicas de los osteíctios[4]
Los distintos tipos de escamas actuales derivan de las que presentaban los ostracodermos o placodermos. En la mayoría de los grupos se suprimió la capa superior, quedándose sólo la parte basal formada por hueso laminar. Esta eliminación sirvió para aligerar el cuerpo y facilitar la natación. Las escamas cosmoideas, gruesas, con hueso laminar, esponjoso, dentina y esmalte, persisten en los sarcopterigios. Las escamas ganoideas son una forma intermedia, con una capa gruesa de hueso laminar, hueso esponjoso y esmalte, presente en algunos grupos, como Lepisosteus.
Los peces modernos tienen las escamas elasmoideas, formadas exclusivamente por hueso laminar. Hay dos tipos de escama elasmoidea: la cicloidea y la ctenoidea. La ctenoidea tiene un borde posterior con unas pequeñas incisiones radiales a modo de peines. La cicloidea tiene un borde posterior liso. Las escamas cicloideas son propias en general de peces más primitivos, y las ctenoideas, de peces más modernos.
Las escamas elasmoideas tienen una estructura en capas circulares. La parte central de la escama es el focus (plural focii), alrededor del cual aparece una serie de anillos de crecimiento (annulii) que sirven para estimar la edad del individuo. También pueden presentar radios (radii) por su borde. La parte anterior (o campo anterior) de la escama está situada por debajo de otras escamas.
 Escamas de la línea lateral del rutilo (Rutilus rutilus).
Escamas de la línea lateral del rutilo (Rutilus rutilus).Las escamas de la línea lateral son diferentes. Tienen unos agujeros para que la línea lateral funcione correctamente. Estas escamas hacen que la línea lateral se marque y resulte relativamente conspicua.
Cromatóforos
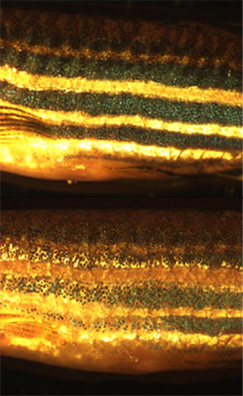
 Los cromatóforos del pez cebra son los encargados de generar el mimetismo con el fondo, al ser expuestos a ambientes oscuros (arriba) o luminosos (abajo).
Los cromatóforos del pez cebra son los encargados de generar el mimetismo con el fondo, al ser expuestos a ambientes oscuros (arriba) o luminosos (abajo).Los cromatóforos de la dermis, responsables de la coloración de la piel, son los melanóforos (contienen melanina, oscurecen la piel y son células muy ramificadas) y los xantóforos o lipóforos (poseen pigmentos de estructura lipídica de color amarillento). La combinación de ambos producen colores rojizos. También existen los iridocitos o guanóforos, que contienen capas de guanina que producen efectos sobre los dos pigmentos mencionados, alterando su coloración. Pueden producir una coloración blanca característica.
Hay peces, como el lenguado o la platija, capaces de imitar la coloración del fondo, en un efecto conocido como metacrosis, o cambio rápido de color. La metacrosis se consigue gracias a la estructura típica de los melanóforos, que permite concentrar la melanina en unas zonas y dispersarla en otras, dando diferentes patrones de coloración. El control de esta capacidad se debe a una vía nerviosa y una vía endocrina. Las hormonas que participan son la adrenalina, que concentra la melanina, y la hormona dispersora de la melanina (MDH, de sus siglas en inglés Melanin-Dispersing Hormone), que la dispersa.
Las coloraciones varían mucho dependiendo de la forma de vida. Los peces epipelágicos son, en su mayor parte, contrasombreados, ya que tienen el lomo oscuro y el abdomen blanco, lo cual sirve para mimetizarse en el medio contra los depredadores (vistos desde una posición inferior, el color blanco del vientre hace que se mimeticen con el reflejo del sol; vistos desde una posición superior, el color oscuro del dorso hace que se mimeticen con el fondo oscuro del mar u océano). Estos mismos peces son plateados por el flanco (el llamado argenteum), también como estrategia de mimetismo. En los peces de río predominan las coloraciones pardas y verdosas, pero todavía hay contrasombreado. Los peces de fondo suelen ser rojos ya que este es el color que primero se pierde a medida que descendemos en profundidad. Además, suelen tener los ojos con un tamaño más grande que los epipelágicos.
Encontramos en ciertas especies coloraciones con diseños disruptivos (como los de las cebras), y coloraciones de despiste, como ocelos o manchas que imitan ojos. Las coloraciones crípticas pueden llegar a ser muy exageradas, y se dan tanto en peces depredadores como en potenciales presas.
Se observan asimismo coloraciones fanéricas (llamativas) que frecuentemente se dan en época de celo, o bien sirven para avisar de que el animal posee mecanismos de defensa especial (coloraciones aposemáticas). Un ejemplo típico de coloración aposemática son los peces payaso (Amphiprion), que viven en los tentáculos de las anémonas. Acanthurus, el pez cirujano, tiene una espina muy cortante para defenderse y avisa con un aspecto muy llamativo. Algunos peces limpiadores tienen coloración fanérica para ser reconocidos como tales; también hay especies imitadoras de éstas últimas, haciéndose pasar por limpiadores y aprovechando la confianza del individuo para alimentarse de él.
-

Contrasombreado en (Thunnus thynnus)
-

Diseño disruptivo en (Dascyllus melanurus)
-

Coloración críptica en (Synanceia verrucosa)
-

Coloración aposemática en (Amphiprioninae)
Fotóforos
Los fotóforos son órganos que emiten luz. Se da en muchas especies batipelágicas. Los fotóforos tienen normalmente una estructura de glándula. Estos órganos de producción de luz varían mucho de unas especies a otras. En algunos casos se transforman las escamas sobre los fotóforos para que actúen de lentes, y otras veces vienen acompañadas de guanóforos. Sirven para reconocer individuos de la misma especie, para localizar presas o para atraerlas. Aparecen en muchos grupos, incluidos en algunos en los condrictios, por lo que es un proceso de convergencia adaptativa.
Sistema esquelético
El esqueleto interno de los osteíctios está calcificado, con la excepción de los esturiones, que es cartilaginoso. Está compuesto por el esqueleto axial (columna vertebral), el esqueleto cefálico (cabeza), el esqueleto zonal (las dos cinturas que se anclan a las aletas pelvianas y torácicas respectivamente) y el esqueleto apendicular (aletas).
 Esqueleto generalizado de un osteíctio.
Esqueleto generalizado de un osteíctio.Esqueleto axial
Está formado por la columna vertebral, que se subdivide a su vez en región troncal (conjunto de vértebras que presentan costillas y carecen de arco hemal) y región caudal (conjunto de vértebras que carecen de costillas y presentan arco hemal). Las vértebras están osificadas y presentan un canal central muy pequeño por el que discurre la notocorda, presente también en los cojinetes situados entre las vértebras. Cada una es cóncava por delante y por detrás, y poseen encima un arco neural (por el que pasa el cordón nervioso) y por debajo un arco hemal (por el que pasa un vaso sanguíneo y presente solo en el esqueleto axial caudal).
Al tipo de vértebras que encontramos en los Condrictios las llamamos anficélicas, y resultan útiles para la natación, confiriendo flexibilidad al esqueleto axial. La notocorda queda comunicada por el centro de cada vértebra, los cojinetes entre vértebra y vértebra no están aislados.
En la región troncal hay dos tipos de costillas: las ventrales y las dorsales. Las ventrales son homólogas a las de los tetrápodos. En el septo horizontal de tejido conjuntivo que separa la musculatura ventral de la dorsal aparece el juego dorsal de costillas.
Esqueleto cefálico
Está formado por el cráneo, y protege el encéfalo, los órganos de los sentidos y los arcos branquiales. En comparación con el cráneo de los tiburones, los osteíctios presentan unos añadidos de esqueleto dérmico óseo (sin precursor cartilaginoso, conocido en conjunto como dermatocráneo, derivado del caparazón de los ostracodermos) por encima del cráneo interno (con precursor cartilaginoso, conocido con conjunto como endocráneo). Al cráneo que protege el encéfalo le damos el nombre de neurocráneo; el resto forma parte del esplacnocráneo.
El cráneo es de gran complejidad, mayor que el de los tetrápodos. En la parte del endocráneo, destacamos el hueso etmoides, el alisfenoides y el esfenoides en la parte más anterior. En la parte de los órganos de oído aparece el hueso ótico, y en la parte que engancha a la columna vertebral están el occipital y el paraoccipital. El dermatocráneo se dispone por encima de estos huesos, dando el hueso nasal, el frontal etc.
El dermatocráneo que forma un techo por arriba se llama bóveda dérmica, y ha sido heredado por los tetrápodos. Por debajo del neurocráneo hay otros huesos: palatinos, pterigoideos o pterigoides, vómer y parasfenoides. Estos huesos forman el paladar. El conjunto de estos huesos junto con los que componen la mandíbula superior (maxila) recibe el nombre de complejo maxilo-palatino. Este complejo deriva de los dos cartílagos que los tiburones tienen en las mandíbulas: el palatocuadrado da por osificación el cuadrado y el complejo palatino-pterigoideo, el cartílago de Meckel da lugar al hueso articular de la mandíbula inferior. No obstante, a estos huesos derivados del cartílago se superponen el maxilar y el premaxilar (se disponen sobre el palatocuadrado), que son huesos que funcionalmente reemplazan al palatocuadrado.
Sobre el cartílago de Meckel (hueso articular) se superpone el dentario, que es otro hueso dérmico. Los dientes en los osteíctios aparecen sobre los huesos dérmicos. El siguiente arco es el hioideo, que colabora en la sujeción de la mandíbula (sujeción hiostílica, como en los tiburones). Aparece un hueso intermedio, el simpléctico, que une el hiomandibular al cuadrado. El hioides viene a ser el esqueleto de la lengua. En los arcos que siguen a partir de éste ya no hay superposición de huesos dérmicos.
Los peces óseos tienen la capacidad de proyectar la boca, lo que permite obtener un efecto de succión. Esta proyección es del premaxilar, maxilar y dentario, que son los que llevan los dientes. Hay otro conjunto de huesos dérmicos, que son los huesos operculares. Estos huesos se disponen por encima del arco hioideo. Son el preopercular, opercular, subopercular e interopercular. Por debajo de la cabeza, cerrando las agallas por esa zona, hay unos radios con una membrana entre ellos, y se conocen como radios branquiósteos.
Esqueleto zonal
El esqueleto zonal está compuesto por las dos cinturas, escapular o pectoral y pélvica, en que se anclan las dos extremidades pares. La cintura escapular es la más anterior, y presenta elementos dérmicos (huesos dérmicos), ausentes en la cintura pélvica.
La cintura escapular, en su parte interna, está formada por la escápula y el coracoides, frecuentemente unidos, englobados por una serie de huesos dérmicos. Existe un hueso dérmico grande, el cleito, al que se sujetan la escápula y el coracoides. Por encima de éste encontramos el supracleito, y más aún por encima otro hueso más, el postemporal. A veces en esta cintura hay una clavícula. La cintura escapular se apoya algo en el cráneo.
La cintura pelviana tiene una pieza impar (el isquiopúbico o pelvis, procedente de dos piezas que se fusionaron). Esta cintura no se apoya en el cráneo, y gracias a otra pieza suelta, el basipterigio, da flexibilidad a las extremidades.
Esqueleto apendicular
El esqueleto apendicular de los peces está formado por las aletas. En las aletas impares podemos distinguir dos zonas: el pterigóforo y los radios. El pterigóforo es la parte del esqueleto apendicular que se inserta en el animal internamente, en un septo de tejido conjuntivo. Los radios son la continuación en el exterior del animal de los pterigóforos, y pueden ser simples (de una sola pieza) o blandos (de varias piezas articuladas pudiendo estar ramificadas). Podrían derivarse de escamas, por lo que se clasifican como lepidotricos.
En las aletas pares hay también unas piezas basales y unos radios. En los elementos basales la estructura es muy variable. En las especies más primitivas son piezas grandes y complejas. En los teleósteos estas piezas han tendido a perder tamaño y aumentar en número.
La aleta caudal es en la gran mayoría de los casos homocerca (en los esturiones es heterocerca). Para conseguir la cola homocerca se ha desarrollado una pieza oblicua, denominada urostilo, que se interpreta que se trata de restos de centros vertebrales de la cola heterocerca que se han fusionado. La forma externa homocerca se ha obtenido por la formación de siete huesos hipurales derivados de los radios de la cola heterocerca. Por lo tanto, la simetría de la cola homocerca es sólo exterior, puesto que la estructura interna es muy asimétrica.
Sistema muscular
La musculatura completa de los osteíctios está dividida, básicamente, en cuatro partes resultado de dos divisiones. La primera división o tabicación se debe a un septo axial o longitudinal, que separa una mitad izquierda y otra derecha. La segunda división se debe a un septo horizontal o transverso, donde se sitúan las costillas dorsales, y lo dividen en el músculo epiaxial (el de arriba) y el músculo hipoaxial (el de abajo).
Los paquetes musculares se apoyan cada uno en el siguiente para incrementar la eficacia del movimiento. Esta disposición curva de los septos y los paquetes musculares es la que da a la carne de los peces su aspecto característico.
La musculatura apendicular deriva de la axial, y se da sólo en la base de la aleta. La musculatura cefálica sirve para abrir y cerrar la boca y controlar el hioides.
En muchas especies existe musculatura eléctrica. Los Monorrínidos, por ejemplo, se comunican por electricidad y la utilizan para percibir obstáculos. Cada especie de este grupo tiene una frecuencia de ondas eléctricas características, siendo éste uno de los mecanismos usados para identificar individuos de su misma especie. Ejemplos de esta musculatura son Electrophorus electricus con musculatura eléctrica esquelética, y Uroscopus con musculatura eléctrica derivada del músculo ocular.
Sistema digestivo
 Posiciones de la boca en los peces:
Posiciones de la boca en los peces:
(a) - Terminal,
(b) - Superior,
(c) - Subterminal, inferiorEl sistema digestivo consta de un tubo con una serie de glándulas anejas. En el inicio está la cavidad bucal, y después una faringe donde se localizan las hendiduras branquiales. La boca es muy variable en tamaño y forma, pudiendo ser ínfera, terminal subterminal u orientada hacia arriba. Presentan un estómago normal, en forma de U o carecen de él. Cuando existe, al final pueden aparecer unos ciegos pilóricos.
Los dientes son muy variables. Los típicos son pequeños, cónicos y dispuestos en el borde de la mandíbula (acrodontos), sujetos con tejido conjuntivo (ligamentos), lo que les permite cierta movilidad, útil en la bascularización hacia dentro y permanencia en una posición recta y fija para la captura de presas. A veces encontramos dientes en los arcos branquiales, y a veces aparece heterodoncia, es decir, dientes con diferentes formas.
Las glándulas anejas al tubo digestivo más importantes son el hígado, que es rico en vitaminas A y D y el bazo.
Dieta
El tipo de dieta condiciona la dentición y la forma general del aparato digestivo. Lo más frecuente es la dieta carnívora, en detrimento de la dieta herbívora. Los carnívoros tienen el estómago grande y dilatable, con bastantes ciegos pilóricos e intestino corto y recto. En los herbívoros el estómago es simple, a veces con una parte diferenciada en estómago triturador, con intestino largo y complejo. Los peces que se alimentan de plancton poseen unos dientes que han reducido su tamaño o han desaparecido, ya que el alimento se retiene en los arcos branquiales.
Sistema respiratorio
La respiración en los osteíctios puede ser por branquias o pulmones. La vejiga natatoria (vejiga gaseosa) deriva de la pared dorsal del esófago, y tiene un origen distinto al pulmón, aunque existan casos de peces que usan la vejiga natatoria como pulmón, pero es sólo una convergencia adaptativa. Esta vejiga aparece en posición dorsal, y a veces tiene una conexión directa con el intestino. De hecho la vejiga natatoria se forma como un pequeño divertículo del digestivo. El canal que comunica la vejiga con el digestivo se llama canal pneumático, y permite introducir aire en esta. Este conducto sólo aparece en peces primitivos (peces fisóstomos), y en los más evolucionados está obstruído (obliterado) (peces fisoclistos). El conducto pneumático obtiene su nombre de que lleva aire.
Los Osteíctios actuales tienen cinco arcos branquiales. El septo correspondiente al arco hioideo es el que aloja a los huesos operculares. Las branquias tienen un sistema de contracorriente, lo que aumenta la efectividad del intercambio gaseoso (captación de oxígeno). El espiráculo desaparece, salvo en los esturiones. Los septos intrabranquiales (salvo el opérculo) entran en regresión (en el esturión y en los Holósteos está a la mitad) y terminan desapareciendo en los más modernos (Teleósteos: tienen dos láminas branquiales por arco branquial de cada lado, no separadas ya por ningún septo).
Un pez Teleósteo tiene cuatro branquias dobles a cada lado. Cada una de estas branquias es doble porque tiene dos láminas no separadas por un septo.
Hacia el interior de la boca aparecen unas espinas (branquispinas), muy desarrolladas en los filtradores. También se encuentran dientes faríngeos que se forman en los arcos branquiales.
La vejiga gaseosa:
En los más modernos tiende a diferenciar dos partes: la cámara oval (más adelantada), separada por un esfínter muscular del resto de la vejiga. El esfínter sirve para poder vaciar rápidamente la vejiga.Los fisoclistos llenan la vejiga segregando gas a su interior en los cuerpos rojos, que son regiones con una rete mirabile de capilares. Se secreta oxígeno a la vejiga, obtenido de la transformación de glucosa en ácido láctico. Para vaciar la vejiga interviene la cámara oval: se abre el esfínter, entra el aire en la cámara oval, y en ésta (que tiene las paredes muy finas) el oxígeno pasa a la sangre.
Los pulmones:
Son ventrales. La vejiga deriva evolutivamente de los pulmones. El pulmón aparece por ejemplo en los Dipnoos, como Protopterus (peces pulmonados africanos).El pulmón se comunica con el esófago mediante una pequeña tráquea. Tiene alveolos, hasta tres generaciones (un alveolo, con alveolos que a su vez tienen alveolos).
La derivación de los pulmones a la vejiga natatoria comenzaría con la reducción de su número a uno solo, y el desplazamiento de éste a la región dorsal (lo que permite que quede por encima del punto de equilibrio). Tras esto ya aparecería el esfínter y desaparecería el conducto pneumático.
Muchos peces emplean la vejiga natatoria como un sistema complementario para la obtención de oxígeno. Se plantea que los pulmones podrían haber aparecido antes de la separación entre Condrictios y Osteíctios, pero en los Condrictios habría desaparecido por completo.La vejiga tiene a menudo otras funciones:
Puede servir para producir sonidos o para recibirlos. Algunas especies hacen vibrar arcos de las vértebras para hacer ruidos que son amplificados por la vejiga (que actúa como una caja de resonancia). Otros utilizan las branquispinas de los arcos branquiales, que hacen chocar para producir sonido.Cuando la vejiga sirve para percibir sonidos, los dos oídos internos comunican mediante osículos (los osículos de Weber) con la vejiga. De modo que el aparato queda como: oído interno-saco endolinfático-osículos de Weber-vejiga natatoria. Parece ser que los osículos de Weber derivan de vértebras.
Reproducción
La reproducción es generalmente ovípara, con puestas que pueden llegar a ser de millones de huevos, con objeto de compensar la alta mortalidad que sufren las crías. Ocasionalmente también se da la reproducción ovovivípara y vivípara. La fecundación es externa, por ello, al contrario de lo que ocurre en los condrictios, los machos no poseen órganos copuladores. Los hábitos reproductores son muy sofisticados, incluso con actividades como construcción de nidos o cuidado de las crías. Los sexos pueden ser separados, hermafroditas, e incluso existir intercambio de sexos en determinadas etapas de la vida. En algunas especies de peces abisales existe dimorfismo sexual (el macho es mucho más pequeño que la hembra, y vive como parásito pegado a ella durante toda su vida).
Véase también
- Evolución de los peces.
Referencias
- ↑ The Taxonomicon - Osteichthyes
- ↑ Froese, R. & Pauly, D. (Editors). 2009. FishBase. World Wide Web electronic publication. www.fishbase.org, version (10/2009).
- ↑ Tree of Life - Gnathostomata
- ↑ Varios autores (2005). «Peces». Animal. La definitiva e impactante guía visual de la vida salvaje en nuestro planeta. Madrid: Pearson Educación, S.A.. ISBN 84-205-3616-4.
Enlaces externos
 Wikiespecies tiene un artículo sobre Osteichthyes. Wikispecies
Wikiespecies tiene un artículo sobre Osteichthyes. Wikispecies Wikimedia Commons alberga contenido multimedia sobre Osteichthyes. Commons
Wikimedia Commons alberga contenido multimedia sobre Osteichthyes. Commons
Categorías:- Fósiles temporal
- Osteichthyes












Wikimedia foundation. 2010.