- Leyes de Mendel
-
Las Leyes de Mendel son un conjunto de reglas básicas sobre la transmisión por herencia de las características de los organismos padres a sus hijos. Estas reglas básicas de herencia constituyen el fundamento de la genética. Las leyes se derivan del trabajo realizado por Gregor Mendel publicado en el año 1865 y el 1866, aunque fue ignorado por largo tiempo hasta su redescubrimiento en 1900.
La historia de la ciencia encuentra en la herencia mendeliana un hito en la evolución de la biología sólo comparable con las Leyes de Newton en el desarrollo de la Física. Tal valoración se basa en el hecho de que Mendel fue el primero en formular con total precisión una nueva teoría de la herencia, expresada en lo que luego se llamaría "Leyes de Mendel", que se enfrentaba a la poco rigurosa teoría de la herencia por mezcla de sangre. Esta teoría aportó a los estudios biológicos las nociones básicas de la genética moderna.[1]
No obstante, no fue sólo su trabajo teórico lo que brindó a Mendel su envergadura científica a los ojos de la posteridad; no menos notables han sido los aspectos epistemológicos y metodológicos de su investigación. El reconocimiento de la importancia de una experimentación rigurosa y sistemática, y la expresión de los resultados observacionales en forma cuantitativa mediante el recurso a la estadística ponían de manifiesto una postura epistemológica totalmente novedosa para la biología de la época.[2] Por esta razón, la figura de Mendel suele ser concebida como el ejemplo paradigmático del científico que, a partir de la meticulosa observación libre de prejuicios, logra inferir inductivamente sus leyes, que en el futuro constituirían los fundamentos de la genética. De este modo se ha integrado el trabajo de Mendel a la enseñanza de la biología: en los textos, la teoría mendeliana aparece constituida por las famosas dos leyes, concebidas como generalizaciones inductivas a partir de los datos recogidos a través de la experimentación.[3]
Historia
La teoría de la herencia por mezcla suponía que los caracteres se transmiten de padres a hijos mediante fluidos corporales que, una vez mezclados, no se pueden separar, de modo que los descendientes tendrán unos caracteres que serán la mezcla de los caracteres de los padres. Esta teoría, denominada pangénesis, se basaba en hechos tales como que el cruce de plantas de flores rojas con plantas de flores blancas producen plantas de flores rosas. La pangénesis fue defendida por Anaxágoras, Demócrito y los tratados hipocráticos y, con algunas modificaciones, por el propio Charles Darwin.
Las leyes de Mendel de la herencia fueron derivadas de las investigaciones sobre cruces entre plantas realizadas por Gregor Mendel, un monje agustino austriaco, en el siglo XIX. Entre los años 1856 y 1863,Gregor Mendel cultivó y probó cerca de 28.000 plantas de la especie Pisum sativum (guisante). Sus experimentos le llevaron a concebir dos generalizaciones que después serían conocidas como Leyes de Mendel de la herencia o herencia mendeliana. Las conclusiones se encuentran descritas en su artículo titulado Experimentos sobre hibridación de plantas (cuya versión original en alemán se denomina “Versuche über Plflanzenhybriden”) que fue leído a la Sociedad de Historia Natural de Brno el 8 de febrero y el 8 de marzo de 1865 y posteriormente publicado en 1866.[4]
 Gregor Johann Mendel descubridor de las leyes básicas de la herencia biológica.
Gregor Johann Mendel descubridor de las leyes básicas de la herencia biológica.
Mendel envió su trabajo al botánico suizo Karl von Nägeli (una de las máximas autoridades de la época en el campo de la biología). Fue él quien le sugirió que realizara su serie de experimentos en varias especies del género Hieracium. Mendel no pudo replicar sus resultados, ya que posteriormente a su muerte, en 1903, se descubrió que en Hieracium se producía un tipo especial de partenogénesis, provocando desviaciones en las proporciones mendelianas esperadas. De su experimento con Hieracium, Mendel posiblemente llegó a pensar que sus leyes sólo podían ser aplicadas a ciertos tipos de especies y, debido a esto, se apartó de la ciencia y se dedicó a la administración del monasterio del cuál era monje. Murió en 1884, completamente ignorado por el mundo científico.
En 1900, sin embargo, el trabajo de Mendel fue redescubierto por tres científicos europeos, el holandés Hugo de Vries, el alemán Carl Correns, y el austríaco Erich von Tschermak, por separado, y sin conocer los trabajos de Mendel llegaron a las mismas conclusiones que él. De Vries fue el primero que publicó sobre las leyes, y Correns, tras haber leído su artículo y haber buscado en la bibliografía publicada, en la que encontró el olvidado artículo de Mendel, declaró que éste se había adelantado y que el trabajo de De Vries no era original. En realidad, la idea de que los factores eran partículas físicas no se impondría hasta principios del siglo XX. Parece más probable que Mendel interpretó los factores de herencia en términos de la filosofía neoaristotélica, interpretando las características recesivas como potencialidades y las dominantes como actualizaciones[5]
En Europa fue William Bateson, quien impulsó en 1900 el conocimiento de las leyes de Mendel. Al dar una conferencia en la Sociedad de Horticultura, tuvo conocimiento del trabajo de Mendel, a través del relato de Hugo de Vries; así encontró el refrendo de lo que había estado experimentando. Él fue, pues, quien dio las primeras noticias en Inglaterra de las investigaciones de Mendel. En 1902, publicó “Los principios mendelianos de la herencia”: una defensa acompañada de la traducción de los trabajos originales de Mendel sobre hibridación. Además, fue el primero en acuñar términos como "genética", "gen" y "alelo" para describir muchos de los resultados de esta nueva ciencia biológica.
En 1902, Theodore Boveri y Walter Sutton, trabajando de manera independiente, llegaron a una misma conclusión y propusieron una base biológica para los principios mendelianos, denominada “Teoría cromosómica de la herencia”. Esta teoría sostiene que los genes se encuentran en los cromosomas y al lugar cromosómico ocupado por un gen se le denominó locus (se habla de loci si se hace referencia al lugar del cromosoma ocupado por varios genes). Ambos se percataron de que la segregación de los factores mendelianos (alelos) se correspondía con la segregación de los cromosomas durante la división meiótica (por tanto, existía un paralelismo entre cromosomas y genes).
Algunos trabajos posteriores de biólogos y estadísticos tales como R.A. Fisher (1911) mostraron que los experimentos realizados por Mendel tenían globalidad en todas las especies, mostrando ejemplos concretos de la naturaleza. Los principios de la segregación equitativa (2ª ley de Mendel) y la transmisión independiente de la herencia (3ª ley de Mendel) derivan de la observación de la progenie de cruzamientos genéticos, no obstante, Mendel no conocía los procesos biológicos que producían esos fenómenos.
Así, puede considerarse que las leyes de Mendel reflejan el comportamiento cromosómico durante la meiosis: la primera ley responde a la migración aleatoria de los cromosomas homólogos a polos opuestos durante la anafase I de la meiosis (tanto los alelos como los cromosomas homólogos segregan de manera equitativa o 1:1 en los gametos) y la segunda ley, al alineamiento aleatorio de cada par de cromosomas homólogos durante la metafase I de la meiosis (por lo que genes distintos y pares diferentes de cromosomas homólogos segregan independientemente).
Experimentos
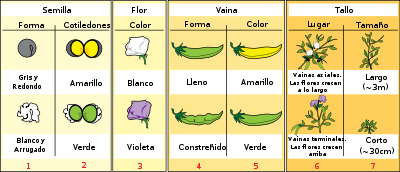 Los siete caracteres que observó G. Mendel en sus experiencias genéticas con los guisantes.
Los siete caracteres que observó G. Mendel en sus experiencias genéticas con los guisantes.Mendel publicó sus experimentos con guisantes en 1865 y 1866. A continuación se describen las principales ventajas de la elección de Pisum sativum como organismo modelo: su bajo coste, tiempo de generación corto, elevado índice de descendencia, diversas variedades dentro de la misma especie (color, forma, tamaño, etc.). Además, reúne características típicas de las plantas experimentales, como poseer caracteres diferenciales constantes.
Pisum sativum es una planta autógama, es decir, se autofecunda. Mendel lo evitó emasculándola (eliminando las anteras). Así pudo cruzar exclusivamente las variedades deseadas. También embolsó las flores para proteger a los híbridos de polen no controlado durante la floración. Llevó a cabo un experimento control realizando cruzamientos durante dos generaciones sucesivas mediante autofecundación para obtener líneas puras para cada carácter.
Mendel llevó a cabo la misma serie de cruzamientos en todos sus experimentos. Cruzó dos variedades o líneas puras diferentes respecto de uno o más caracteres. Como resultado obtenía la primera generación filial (F1), en la cuál observó la uniformidad fenotípica de los híbridos. Posteriormente, la autofecundación de los híbridos de F1 dio lugar a la segunda generación filial (F2), y así sucesivamente. También realizó cruzamientos recíprocos, es decir, alternaba los fenotipos de las plantas parentales:
♀P1 x ♂P2♀P2 x ♂P1
(siendo P la generación parental y los subíndices 1 y 2 los diferentes fenotipos de ésta).
Además, llevó a cabo retrocruzamientos, que consisten en el cruzamiento de los híbridos de la primera generación filial (F1) por los dos parentales utilizados, en las dos direcciones posibles:
♀F1 x ♂P2 y ♀P2 x ♂F1 (cruzamientos recíprocos)♀F1 x ♂P1 y ♀P1 x ♂F1 (cruzamientos recíprocos)
Los experimentos demostraron que:
- La herencia se transmite por elementos particulados (refutando, por tanto, la herencia de las mezclas).
- Siguen normas estadísticas sencillas, resumidas en sus dos principios.
Las leyes de Mendel
Las tres leyes de Mendel explican y predicen cómo van a ser los caracteres físicos (fenotipo) de un nuevo individuo. Frecuentemente se han descrito como «leyes para explicar la transmisión de caracteres» (herencia genética) a la descendencia. Desde este punto de vista, de transmisión de caracteres, estrictamente hablando no correspondería considerar la primera ley de Mendel (Ley de la uniformidad). Es un error muy extendido suponer que la uniformidad de los híbridos que Mendel observó en sus experimentos es una ley de transmisión, pero la dominancia nada tiene que ver con la transmisión, sino con la expresión del genotipo. Por lo que esta observación mendeliana en ocasiones no se considera una ley de Mendel. Así pues, hay tres leyes de Mendel que explican los caracteres de la descendencia de dos individuos, pero solo son dos las leyes mendelianas de transmisión: la Ley de segregación de caracteres independientes (2ª ley, que, si no se tiene en cuenta la ley de uniformidad, es descrita como 1ª Ley) y la Ley de la herencia independiente de caracteres (3ª ley, en ocasiones descrita como 2ª Ley).
1ª Ley de Mendel: Ley de la uniformidad
Establece que si se cruzan dos razas puras para un determinado carácter, los descendientes de la primera generación serán todos iguales entre sí fenotipica y genotipicamente e iguales fenotipicamente a uno de los progenitores..
2ª Ley de Mendel: Ley de la segregación
Conocida también, en ocasiones como la primera Ley de Mendel, de la segregación equitativa o disyunción de los alelos. Esta ley establece que durante la formación de los gametos y cada alelo de un par se separa del otro miembro para determinar la constitución genética del gameto filial. Es muy habitual representar las posibilidades de hibridación mediante un cuadro de Punnett.
Mendel obtuvo esta ley al cruzar diferentes variedades de individuos heterocigotos (diploides con dos variantes alélicas del mismo gen: Aa), y pudo observar en sus experimentos que obtenía muchos guisantes con características de piel amarilla y otros (menos) con características de piel verde, comprobó que la proporción era de 3:4 de color amarilla y 1:4 de color verde (3:1).
Según la interpretación actual, los dos alelos, que codifican para cada característica, son segregados durante la producción de gametos mediante una división celular meiótica. Esto significa que cada gameto va a contener un solo alelo para cada gen. Lo cual permite que los alelos materno y paterno se combinen en el descendiente, asegurando la variación.
Para cada característica, un organismo hereda dos alelos, uno de cada pariente. Esto significa que en las células somáticas, un alelo proviene de la madre y otro del padre. Éstos pueden ser homocigotos o heterocigotos.
En palabras del propio Mendel:[6]
"Resulta ahora claro que los híbridos forman semillas que tienen el uno o el otro de los dos caracteres diferenciales, y de éstos la mitad vuelven a desarrollar la forma híbrida, mientras que la otra mitad produce plantas que permanecen constantes y reciben el carácter dominante o el recesivo en igual número. "
Gregor Mendel3ª Ley de Mendel: Ley de la recombinación independiente de los factores
En ocasiones es descrita como la 2ª Ley. Mendel concluyó que diferentes rasgos son heredados independientemente unos de otros, no existe relación entre ellos, por lo tanto el patrón de herencia de un rasgo no afectará al patrón de herencia de otro. Sólo se cumple en aquellos genes que no están ligados (en diferentes cromosomas) o que están en regiones muy separadas del mismo cromosoma. Es decir, siguen las proporciones 9:3:3:1.
En palabras del propio Mendel:[6]
Por tanto, no hay duda de que a todos los caracteres que intervinieron en los experimentos se aplica el principio de que la descendencia de los híbridos en que se combinan varios caracteres esenciales diferentes, presenta los términos de una serie de combinaciones, que resulta de la reunión de las series de desarrollo de cada pareja de caracteres diferenciales.
Gregor MendelPatrones de herencia mendeliana
Mendel describió dos tipos de "factores" (genes) de acuerdo a su expresión fenotípica en la descendencia, los dominantes y los recesivos, pero existe otro factor a tener en cuenta en organismos dioicos y es el hecho de que los individuos de sexo femenino tienen dos cromosomas X (XX) mientras los masculinos tienen un cromosoma X y uno Y (XY), con lo cual quedan conformados cuatro modos o "patrones" según los cuales se puede trasmitir una mutación simple:
- Gen dominante ubicado en un autosoma (herencia autosómica dominante).
- Gen recesivo ubicado en un autosoma (herencia autosómica recesiva).
- Gen dominante situado en el cromosoma X (herencia dominante ligada al cromosoma X).
- Gen recesivo situado en el cromosoma X (herencia recesiva ligada al cromosoma X).
Fenómenos que alteran las segregaciones mendelianas
Herencia ligada al sexo
Es la herencia con el par sexual. El cromosoma X porta numerosos genes en tanto el cromosoma Y tan solo unos pocos y la mayoría en relación con la masculinidad. El cromosoma X es común para ambos sexos, pero solo el hombre posee cromosoma Y.
Herencias influidas por el sexo y limitadas al sexo
En las herencias limitadas al sexo pueden estar comprometidos mutaciones de genes con cromosomas autosómicos cuya expresión solamente tiene lugar en órganos del aparato reproductor masculino o femenino. Un ejemplo es el defecto congénito septum vaginal transverso, de herencia autosómica recesiva, o la deficiencia de 5 α reductasa que convierte a la testosterona en dihidrotestosterona que actúa en la diferenciación de los genitales externos masculinos, por lo que su ausencia simula genitales femeninos cuando el niño nace.
Una mutación puede estar influida por el sexo, esto puede deberse al efecto del metabolismo endocrino que diferencia a machos y hembras. Por ejemplo, en humanos la calvicie se debe al efecto de un gen que se expresa como autosómico dominante, sin embargo en una familia con la segregación de este gen solo los hombres padecen de calvicie y las mujeres tendrán su cabello más escaso después de la menopausia. Otro ejemplo puede ser la deficiencia de la enzima 21 hidroxilasa que interviene en el metabolismo de los glucocorticoides. Cuando esta enzima está ausente, la síntesis de glucocorticoides se desplaza hacia la formación de testosterona y esta hormona está comprometida en la embriogénesis de los genitales externos del varón, por lo que su presencia anormal en el desarrollo de un feto femenino produce la masculinización de los genitales femeninos, mientras que en el caso de un feto varón, solo incrementa el desarrollo de los masculinos. Una anormalidad de este tipo, permitirá sospechar un diagnostico clínico más rápidamente en una niña, basado en el examen de los genitales del recién nacido, que en un niño.
Estructura génica del cromosoma Y
Por tener un solo cromosoma X, a los individuos de sexo masculino no se les pueden aplicar los términos "homocigoto" o "heterocigoto" para genes ubicados en este cromosoma y ausentes en el cromosoma Y. Ya sean genes que expresen el carácter dominante o recesivo, si están situados en el cromosoma X, los varones siempre lo expresarán y al individuo que lo porta se le denomina hemicigoto.
De lo anterior se deduce que, puesto que las hembras tienen un solo tipo de cromosoma sexual, el X, sus gametos siempre tendrán la dotación cromosómica 23,X, mientras los masculinos pueden portar una X, dando lugar a un individuo femenino (XX), o una Y, con lo que se originaría un individuo masculino (XY). Debido a esto se dice que las mujeres son homogaméticas (todos sus gametos tienen igual constitución) y que los hombres son heterogaméticos (tienen gametos 23,X y 23,Y).
Sistema de compensación de dosis génica del cromosoma X
En insectos, tal como se ha visto en Drosophila, se descubrió la existencia de un gen que ejerce de compensador de dosis, cuando se encuentra en dosis única (como ocurre en machos) produce la activación de la expresión de los genes del cromosoma X. En mamíferos no se ha encontrado un gen con función equivalente.
Lionización
La lionización o inactivacion del cromosoma X se produce porque, a diferencia del cromosoma Y, el X tiene gran cantidad de genes activos que codifican para importantes productos, tales como el factor VIII de la coagulación. Podría pensarse, por tanto, que si las hembras tienen dos X deben tener el doble de los productos o enzimas cuyos genes están en ese cromosoma con relación a los individuos del sexo masculino, sin embargo, esto no ocurre así.
Se ha observado en mamíferos que en las células somáticas del sexo femenino (46,XX), solo uno de los dos cromosomas X es activo. El otro permanece inactivo y aparece en células en interfase como un cuerpo denso fuertemente coloreado, que se inactiva y se adosa a la membrana nuclear en la periferia del núcleo, y que recibe el nombre de cuerpo de Barr. La inactivación del cromosoma X tiene lugar en el estado de mórula, alrededor del tercer día después de la fertilización y se completa, en la masa de células internas que darán origen al embrión, al final de la primera semana de desarrollo embrionario. La selección del cromosoma X que se inactivará, es un fenómeno generalmente aleatorio teniendo en cuenta que al ocurrir la fecundación cada cromosoma X tiene origen materno y paterno, en unas células se inactivará el X materno (Xm) y en otras el X paterno (Xp). Una vez que se inactiva uno de los dos cromosomas X las células descendientes mantendrán el mismo cromosoma X inactivo originándose un clon celular (Xm) o (Xp) activos. Es decir, al inicio de la inactivación, ésta es al azar, primero se inactiva al azar cualquiera de las dos X, ya sea la heredada de la madre o del padre; pero una vez ocurrida se mantiene el mismo cromosoma X que se inactivó en la primera célula del clon y las células que deriven de ésta durante el proceso de crecimiento y desarrollo mantendrán en adelante inactivado el mismo cromosoma X.
La inactivación (desactivación) del cromosoma X está determinada por el gen XIST. Este gen esta involucrado en la transcripción específica de inactivación que funciona por un mecanismo de metilación preferencial, esto significa que si no hay ninguna alteración de estructura en los dos cromosomas X del genoma femenino, la inactivación debe ocurrir de forma aleatoria, pero si existiera alguna alteración con gran compromiso en la función de uno de los dos cromosomas X habría una activación no completamente aleatoria. El locus del gen XIST se encuentra localizado en Xq13.3.
La inactivación del X determina consecuencias genéticas y clínicas:
- Compensación de dosis: iguala la dosis de productos de genes con el hemicigótico para genes localizados en el cromososa X, determinando concentraciones proteicas similares en ambos sexos, para genes ligados al X.
- Variaciones en la expresión de mutaciones en hembras heterocigóticas: por ejemplo, presencia de síntomas más o menos severos en hembras portadoras para hemofilias A o B, distrofia muscular Duchenne, distrofias retinianas recesivas ligadas al X.
- Los órganos femeninos se comportan como mosaicos. Este fenómeno se puede manifestar en zonas en las que se manifieste un alelo (procedente del X de la madre) y otras zonas en las que se manifiesta el otro alelo. Se observa en fenómenos como el color del pelaje de algunas hembras de felinos, de forma que los felinos de tres colores son hembras, y los de dos colores son machos;[7] en el albinismo ocular recesivo ligado al X; o en el test inmunohistoquímico para la detección de la distrofina en hembras heterocigóticas para la distrofia muscular Duchenne.
Penetrancia de un gen o de una mutación específica
Penetrancia es el término que se emplea para referirse a la expresión en términos de todo o nada dentro de una población de individuos. Si la mutación se expresa en menos del 100% de los individuos portadores o heterocigóticos se dice que la mutación tiene una penetrancia reducida y que ese individuo aparentemente “sano” para el carácter o enfermedad que se estudia en la familia puede trasmitir la mutación a su descendencia y éstos expresar el defecto. La penetrancia reducida parece ser el efecto de la relación de la mutación en cuestión y otros genes del genoma, con los cuales se encuentra interactuando.
Expresividad de un gen o mutación específica
Expresividad se usa para referirse al grado de severidad que se manifiesta en el fenotipo. En términos clínicos, es sinónimo de gravedad. La expresión de un gen también depende de la relación de éste con el resto del genoma, pero también de la relación genoma-ambiente. Para referirse a estas gradaciones fenotípicas se utiliza el término expresividad variable del gen o de la mutación.
Efecto pleiotrópico de un gen o mutación específica
Con en término pleiotropía o efecto pleiotrópico de un gen se hace referencia a todas las manifestaciones fenotípicas en diferentes órganos o sistemas que son explicables por una simple mutación. Un ejemplo clásico para explicar este término lo constituye el síndrome Marfan, cuya mutación afecta al gen FBN1 que codifica a la proteína fibrilina, esta proteína se encuentra en el tejido conectivo y explica las manifestaciones esqueléticas, oculares y cardiovasculares que caracterizan al síndrome.
Heterogeneidad genética
Este término se aplica tanto a mutaciones en genes localizados en diferentes cromosomas que producen expresión similar en el fenotipo (heterogeneidad no alélica) como a mutaciones que afectan a diferentes sitios del mismo gen (heterogeneidad alélica). Esta categoría complica extraordinariamente el estudio etiológico de variantes del desarrollo de origen genético y constituye una amplia y fundamental fuente de diversidad genética del desarrollo.
Nuevas mutaciones con expresión dominante
Cuando tiene lugar una mutación de novo que se expresa como dominante, o sea, en un genotipo heterocigótico, ocurre que padres que no presentan el efecto de la mutación pueden tener un descendiente afectado. La ausencia de antecedentes familiares, una vez que se excluyen fenómenos como la penetrancia reducida del gen y variaciones mínimas de la expresividad dificulta llegar al planteamiento de una mutación de novo cuando en la literatura el defecto o enfermedad no ha sido reportada con anterioridad, con un tipo específico de herencia.
Efecto de letalidad en un genotipo específico
Algunas mutaciones se expresan de forma tan severa que producen letalidad en un genotipo específico. Un ejemplo pudiera ser el efecto de una doble dosis de una mutación que se expresa como dominante o el efecto en un genotipo hemicigótico, como ocurre en la incontinencia pigmenti, enfermedad humana dominante ligada al cromosoma X.
Herencia en mamíferos
El árbol genealógico
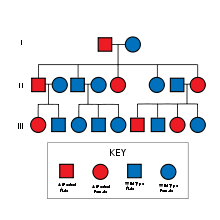 Pedigree autosómico dominante.
Pedigree autosómico dominante.Como en cualquier otra especialidad médica, en genética adquiere enorme importancia el interrogatorio del individuo enfermo y sus familiares, pero, adicionalmente, es vital establecer los lazos de parentesco entre los individuos afectados y los supuestamente sanos, por eso se utiliza el llamado árbol genealógico o pedigree en el que mediante símbolos internacionalmente reconocidos se describe la composición de una familia, los individuos sanos y enfermos, así como el número de abortos, fallecidos, etc.
Herencias dominantes
Cuando el gen productor de una determinada característica (o enfermedad) se expresa aún estando en una sola dosis se denomina dominante y los linajes donde se segrega muestran un árbol genealógico en que, como regla, hay varios individuos que lo expresan y los afectados tienen un progenitor igualmente afectado. No obstante, hay diferencias de acuerdo a si el gen está ubicado en un autosoma o en el cromosoma X.
En la herencia autosómica dominante se cumplen los siguientes hechos:
- Varios individuos afectados.
- Los afectados son hijos de afectados.
- Se afectan por igual hombres y mujeres.
- Como regla, la mitad de la descendencia de un afectado hereda la afección.
- Los individuos sanos tienen hijos sanos.
- Hay hombres afectados hijos de hombres afectados (lo cual excluye la posibilidad de que el gen causante de la afección está ubicado en el cromosoma X, que en los varones procede de la madre).
- El patrón ofrece un aspecto vertical.
En este caso los individuos afectados son usualmente heterocigóticos y tienen un riesgo del 50% en cada intento reproductivo de que su hijo herede la afección independientemente de su sexo.
En la herencia dominante ligada al cromosoma X, aunque el gen sea dominante, si está ubicado en el cromosoma X, el árbol genealógico suele mostrar algunas diferencias con respecto al de la herencia autosómica dominante:
- Aunque los afectados usualmente son hijos de afectados y la mitad de la descendencia presenta la afección, no podemos identificar varones que hayan heredado la afección de su padre, o sea, no hay trasmisión varón-varón, puesto que los padres dan a sus hijos el cromosoma Y.
- Igualmente llama la atención que hay un predominio de mujeres afectadas pues mientras estas pueden heredar el gen de su madre o de su padre, los varones sólo lo adquieren de su madre.
- Una mujer afectada tendrá el 50% de su descendencia afectada, mientras que el hombre tendrá 100% de hijas afectadas y ningún hijo afectado.
Herencias recesivas
Cuando el gen causante de la afección es recesivo, por regla general el número de afectados es mucho menor y suele limitarse a la descendencia de una pareja, pero es más evidente la diferencia en la trasmisión según la mutación esté situada en un autosoma o en el cromosoma X.
En la herencia autosómica recesiva llama la atención la aparición de un individuo afectado fruto de dos familias sin antecedentes. Esto ocurre pues ambos padres de este individuo son heterocigóticos para la mutación, la cual, por ser recesiva, no se expresa ya que existe un alelo dominante normal, pero, como estudiamos en las leyes de Mendel, existe un 25% en cada embarazo, de que ambos padres trasmitan el alelo mutado, independientemente del sexo del nuevo individuo. Por aparecer usualmente en la descendencia de un matrimonio, se dice que su patrón es horizontal. Otro aspecto a señalar es que cuando existe consanguinidad, aumenta la probabilidad de aparición de este tipo de afecciones, debido a que ambos padres comparten una parte de su genoma proporcional al grado de parentesco entre ellos.
En la herencia recesiva ligada al cromosoma X es evidente que los individuos afectados son todos del sexo masculino; esto se justifica porque al tener la mujer dos X y ser el gen recesivo, el alelo dominante normal impide su expresión, mientras el varón hemicigótico si tiene la mutación la expresará. También se observa que entre dos varones afectados existe una mujer, que en este caso es portadora de la mutación. La probabilidad de descendencia afectada dependerá del sexo del progenitor que porta la mutación:
- Un hombre enfermo tendrá 100% de hijas portadoras y 100% de hijos sanos.
- Una mujer portadora tendrá 50% de sus hijas portadoras y 50% de hijos varones enfermos.
Conclusiones y comentarios
Es un error muy extendido suponer que la uniformidad de los híbridos que Mendel observó en sus experimentos es una ley de transmisión, pues la dominancia nada tiene que ver con la transmisión, sino con la expresión del genotipo. Por lo que esta observación mendeliana no suele considerarse una ley. Las leyes mendelianas de transmisión son por lo tanto dos: la Ley de segregación de caracteres independientes (1ª ley) y la Ley de la herencia independiente de caracteres (2ª ley).
Recorriendo la web de Wikipedia se puede observar este hecho, por ejemplo, en las versiones inglesa, francesa y portuguesa consideran que las Leyes de Mendel son dos. En cambio, en otras versiones como la catalana, la alemana, la italiana y la vasca siguen considerando la Ley de la Uniformidad como la primera Ley de Mendel, sin ser estrictamente una ley de transmisión de caracteres.
Incluso a nivel docente y bibliográfico sigue permaneciendo vigente esta visión que debería ser aclarada.
Véase también
- Factor mendeliano: El concepto de factor mendeliano fue introducido en 1860 por Mendel, actualmente denominado gen, éste se puede definir como una unidad física y funcional que ocupa una posición específica en el genoma.
- Gen: Es una región de DNA que codifica para RNA.
- Genotipo: factores hereditarios internos de un organismo, sus genes y por extensión su genoma.
- Fenotipo: las cualidades físicas observables en un organismo, incluyendo su morfología, fisiología y conducta a todos los niveles de descripción.
- Alelo: Es cada una de las variantes de un locus. Cada alelo aporta diferentes variaciones al carácter que afecta. En organismos diploides (2n) los alelos de un mismo locus se ubican físicamente en los pares de cromosomas homólogos.
- Locus: Ubicación del gen en un cromosoma. Para un locus puede haber varios alelos posibles. (Plural: LOCI)
- Cariotipo: Composición fotográfica de los pares de cromosomas de una célula, ordenados según un patrón estándar. En un cariotipo encontramos el conjunto de características que permiten reconocer la dotación cromosómica de una célula.
- Línea pura: Es la descendencia de uno o más individuos de constitución genética idéntica, obteniéndose por autofecundación o cruces endogámicos. Son individuos homocigotos para todos sus caracteres.
- Autofecundación: Proceso de reproducción sexual donde los gametos masculinos de un individuo se fecundan con los óvulos del mismo individuo. Es indispensable que sean especies monoicas (característico de las plantas y algunos animales inferiores).
- Dominancia, Alelo dominante: Predominio de la acción en un alelo sobre la de su alternativo (llamado alelo recesivo), enmascarando u ocultando sus efectos. El carácter hereditario dominante es el que se manifiesta en el fenotipo (conjunto de las propiedades manifiestas en un individuo). Según la terminología mendeliana se expresa como A>a (el alelo A domina sobre el alelo a, el carácter que determina, es por tanto el que observaremos en el fenotipo).
- Recesividad, Alelo recesivo: Característica del alelo recesivo de un gen que no se manifiesta cuando está presente el alelo dominante. Para que este alelo se observe en el fenotipo, el organismo debe poseer dos copias del mismo alelo, es decir, debe ser homocigoto para ese gen (según la terminología mendeliana, se expresaría como “aa”).
- Meiosis: La meiosis es el proceso de división celular que permite a una célula diploide generar células haploides en eucariotas. En este proceso se produce una replicación del DNA (en la fase S) y dos segregaciones cromosómicas, de manera que de una célula inicial diploide se obtienen cuatro células haploides.
- Homocigoto: Individuo puro para uno o más caracteres, es decir, que en ambos loci posee el mismo alelo (representado como aa en el caso de ser recesivo o AA si es dominante).
- Heterocigoto: Individuo que para un gen, tiene un alelo distinto en cada cromosoma homólogo. Su representación mendeliana es “Aa”.
- Híbrido: Es el resultado del cruzamiento o apareamiento de dos individuos puros homocigotos (uno de ellos recesivo y el otro dominante) para uno o varios caracteres.
- Gameto: Célula sexual que procede de una estirpe celular llamada línea germinal, en los seres superiores tienen un número de cromosomas haploide (n) debido a un tipo de división celular llamado meiosis que permite reducir el número de cromosomas a la mitad. El gameto femenino se denomina óvulo; el gameto masculino recibe el nombre de espermatozoide.
- Cigoto o huevo: Célula resultante de la unión de dos gametos haploides (es por tanto, diploide, 2n). Generalmente, experimenta una serie de divisiones celulares hasta que se constituye en un organismo completo. Su citoplasma y sus orgánulos son siempre de origen materno al proceder del óvulo.
- Haploide: Que posee un solo juego de cromosomas (n), característico de los gametos eucariotas y los gametofitos de las plantas.
- Diploide: Que tiene doble juego de cromosomas (2n). Características de las células somáticas.
- Autosoma: Todo cromosoma que no sea sexual.
Referencias
- ↑ Blanc M. Gregor Mendel: la leyenda del genio desconocido. Mundo Científico 1984; 4:274-287.
- ↑ Mayr E. The Growth of Biological Thought. Harvard: Belknap Press of Harvard University Press, 1982.
- ↑ Lombardi, O. ¿Leyes de Mendel o Ley de Mendel? Antropología
- ↑ Henig, Robin Marantz (2009). The Monk in the Garden : The Lost and Found Genius of Gregor Mendel, the Father of Genetics. Houghton Mifflin. ISBN 0-395-97765-7. «The article, written by a monk named Gregor Mendel...»
- ↑ Depew y Weber, 1995
- ↑ a b Sinnott, Edmund Ware; Dunn LC & Dobzhansky T. (1961). «Mendel G. Experimentos de hibridación en plantas, 1866.». Principios de Genética.. Barcelona: Omega. pp. 528-549. B 12915-1961.
- ↑ El PaleoFreak. Mosaicos femeninos.
Bibliografía
- Mendel, Gregor. Experiments in plant hybridization. (1865). Read at the February 8th, and March 8th, 1865, meetings of the Brünn Natural History Society (Original en alemán: Mendel, Gregor. 1866. Versuche über Plflanzenhybriden. Verhandlungen des naturforschenden Vereines in Brünn, Bd. IV für das Jahr 1865, Abhandlungen, 3–47.)
- Griffiths, A.J.F.; S.R. Wessler; R.C. Lewontin & S.B. Carrol (2008). Introducción al análisis genético. 9th edición. McGraw-Hill Interamericana.
- Alberts, Bray, Hopkin, Johnson, Lewis, Raff, Roberts, Walter. Introducción a la Biología Celular. Editorial Médica Panamericana.
- [1]
- [2]
Enlaces externos



Wikimedia foundation. 2010.